
▪ Bible Physics ▪ Epinasty ▪ Plant Senescence Theory ▪ Scripture Evolution ▪ Socrates/Plato Civilization Cycle ▪ Speculations of Why Sexes Exist ▪
▪ 1986 Version ▪ 1995 Version ▪ 1999 Version ▪ 2003 Version ▪ 2007 Version a ▪ 2007 Version b ▪ Current Version ▪
Theory
Here I try to put forth what seems to me to be reasonable assumptions about how Plant hormones work.
1. The goal of a plant is to germinate, survive, grow, and reproduce (and either exploit or contribute to life in general – see my summary of a future paper in progress here.
|
|
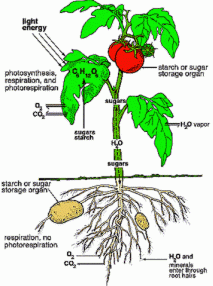
2. The role of the shoot is to create sugars from sunlight, water, and carbon dioxide harvested from the air. It also harvests most of the oxygen needed by the plant for respiration. The shoot may serve as a reserve store for water and minerals. This may be far fetched as a general principle but the storage of water occurs in at least the cactus. The best place for storing all nutrients may be out of harms way in the soil, in the root. The shoot also provides the structure that supports the leaves, flowers, and fruit, but this will not be important here.
3. The role of the root is to harvest water and minerals from the soil. In order to function, the root also needs to harvest some oxygen from spaces between the soil particles. The root also provides a place for storing reserves of sugar in the form of starch and may even store oxygen. It also anchors the plant in a propitious place for it to grow and prevents it from being physically uprooted by the elements or fauna. The anchoring role of the root will not be important here.
4. If they face conditions where they have to make a choice, plants will invest in promising new meristematic cells (i.e. those that are functioning well in their role, like young leaves making good amounts of sugar and successfully harvesting oxygen) and withdraw nutrients from mature cells to feed these “babies,” even if that means withdrawing nutrients from mature cells that are functioning “adequately”. There is a cost to the transfer of nutrients from mature to juvenile cells. This cost is measured in the loss of some nutrients during the process. It is true that minerals cannot be destroyed and are usually not excreted. However, they are probably not fully recoverable from a mature cell, leaf, or root, in the same way that they would be if they were merely stored in some kind internal reserve, such as a vacuole within a cell.
5. There are three general groups of plant hormones. The Growth Hormones are released under long term good growth conditions and are separated into one predominantly synthesized in the shoot and one in the root. The Stress Hormones are released under various kinds of long term stress and are separated into one synthesized predominantly in the shoot and one in the root. Lastly, there are the Shock/Synchronizer Hormones. The idea with the Shock/Synchronizer Hormones is that they are released under rapidly developing stress of any kind or return from stress good conditions, confronting the individual cells, parts of the plant, or the plant as a whole. To elaborate, these are the first hormones released when the physical survival of the cells is under threat or when the cells return to secure environmental and nutrient conditions. They quickly shut a plant or plant part down or restore it to normal functioning. They may also play a secondary role as modulators of the rate of cell metabolism slowing it down to survivable levels according to local conditions, or speeding it up so that full use may be made of current nutrient levels and environmental conditions. In fact, a final climacteric high or sustained level of these hormones may be needed to kick off the synthesis of the stress hormones (GA and Ethylene) on one end, or the growth hormones (Auxin and Cytokinin) on the other.
6. All plant cells are totipotent not just under the right conditions, such as in tissue culture, but also in the way that they behave in response to environmental and nutrient conditions. That is, a shoot cell will always act somewhat like a root cell, and a root cell will always act somewhat like a shoot cell. In addition, a mature cell will act somewhat like an immature cell, and vice versa. For example, below, it is suggested that just like a shoot meristem cell, when any cell is met with good environmental conditions and more than enough sugar and oxygen to support growth, it will make Auxin or at least a tiny amount of it.
7. The Growth Hormones include Auxin and CKs. These are made by all cells when conditions are good for growth. Auxin is made when any plant cell is facing the conditions that would be propitious for the growth of a shoot meristematic cell. These include freedom from environmental stresses and the production or existence of more than enough sugar and oxygen to support it and any cells depending on it. (Root cells, except for the few that are meristematic, have no cells depending on them for their sugar and oxygen needs.) CKs are made by any cell under the conditions that would be propitious for a root meristematic cell to grow. These include freedom from environmental stresses and the uptake or existence of more than enough minerals and water to support it and any dependent cells. To go into more detail, a root cell, for example, would make a CK if it was taking in more than enough minerals and water to support both it and any dependent shoot cell of similar size in the shoot (i.e. a cell in the shoot that depends on that cell in the root for its water and minerals). A similar relationship exists for a shoot cell vis-à-vis sugar and oxygen, and the sister cell in the root that depends on it for the nutrient.
When I say more than enough nutrients for supporting cells, I mean more than enough nutrients to support cells at their peak metabolism. There are two alternatives to this view that I discuss in the addendum.
|
|

8. The Stress Hormones include the GAs, ET and the BRs. These are made by all plant cells under some kind of stress where the plant must remove resources from the site of stress and redistribute the resources to another part of the plant so that the stress is ameliorated. They also initiate the freeing of stored resources to address the particular shortfall. GAs, for instance, are made by all cells under which conditions that would be disadvantageous to a mature root cell (e.g. in any cell when there is less than enough sugar and oxygen to support both it and any dependent cells). GA removes the resources from this cell and redistributes them into some part of the shoot so that the prospect of increasing the supply of sugar and oxygen to the entire plant is increased. (The root would keep the resulting sugar and oxygen for itself and route the minerals and water to where they were needed in the shoot.) Also GA causes the release of enzymes in the root, which turn starch stored in vacuoles into sugar. This sugar also helps, at least temporarily, to resolve deficiencies. I hypothesize that oxygen is also stored in the roots and GA initiates its freeing from storage and availability to the root. Gas is a much less compact stored item than starch, but nevertheless this phenomenon is possible.
Similarly, ET is made by all cells under similar conditions and would be disadvantageous for the existence of a mature shoot cell. For instance, in any cell when there are not enough minerals and water to support both it and any dependent, similar-sized cell, ET causes the withdrawal of all nutrients and redirects the sugar and oxygen to the root, keeping the minerals and water in the shoot. This is an attempt to produce productive new root growth and an eventual surpassing of the previous levels of minerals and water. This is an attempt by the plant at a wise reinvestment of resources. It is a gamble to jump start the growth in mineral and water levels to facilitate the growth of the shoot. These jump starts always have costs. These include some use of energy and thus an overall net loss in the weight of the plant. I hazard that minerals and water are stored in either the shoot (for theory symmetry) or the root (because of the practicality of its inaccessibility to the hostile outside world). I suggest that ET initiates the freeing of stored minerals and water in addition to its resource stress role.
When I say less than enough nutrients for supporting cells, I mean less than enough nutrients to support cells at a survivable or minimum level of metabolism. There are two alternatives to this view that I discuss in the addendum.
Like many involved in the study of plant hormones, I believe that BRs may actually be part of a hormone cascade that involves GA or a parallel path with many similarities. That is, BRs may just be a step along the way in a scheme from a stress to the root cells generally and the plant’s reaction to the stress. In fact, BRs may be the primary hormone igniter of the chemical domino path that leads to the reaction of a plant to stress and GAs may be just a step along the path. Future experimentation will elucidate this. Because of the similarities, I will refer, as some scientists do, to GA and BR together as the GA/BR class of hormones.
9. The Shock/Synchronizer Hormones are the ABAs, and SAs. They fall into two categories: general metabolic inhibitors/senescence stimulators, and general metabolic stimulators/senescence blockers. They act rapidly. I am defining ABAs as general metabolic inhibitors/senescence activators. They would be made when there is any kind of nutrient or environmental stress to a cell, such as a low or high temperature, wounding, mechanical stress from wind or other plants or animals, pests, disease, or the dropping below sustenance level of minerals, water, sugar, or oxygen.
In terms of nutrient stress, the survival ABA is made when any of the resources of the cell fall below sustenance level. This hormone would not be activated if the levels of these nutrients fell below what the particular cell nutrient level was supposed to be for that type of cell if it was supposed to harvest or synthesize that nutrient for others. For instance in a root cell, ABA would not be synthesized until either minerals or water fell below what was necessary for the survival of the root cell itself. The hormone would not be made when the cell had a level of minerals and water below what was necessary to support both it and a sister cell in the shoot. Falling below that level would initiate ET emanation. The level of sugar and oxygen necessary for ABA synthesis in the root would of course be below the sustenance level, which might be a frequent occurrence because the cell has to get these nutrients from the opposite end of the plant. In an attempt to re-establish good supplies GA/BR would also be released when sugar and oxygen fell below sustenance level. Low levels of ABA would be to slow the metabolism so that less sugar and oxygen were needed. At higher ABA levels, ABAs would work with GA/BR to senesce the cell. At low levels, the slowing down of the metabolism itself might retard the GA/BRs’ proposed initiation of that root cell’s senescence. ABA, I hypothesize, always work first at a low level to slow metabolism, so that the nutrient sustenance level is actually lowered to a “bearable level” or in the case of environmental stress of any kind, less damage ensues than would occur at a higher rate of metabolism. When and if the plant or cell “calculates” that survival is not possible or “not warranted,” it releases higher levels of ABA, initiating the senescence of the stressed cell or damaged part of the plant.
The SAs on the other hand are categorized as metabolic stimulators/senescence inhibitors that hasten all metabolic activities and oppose the action of ABA and the Stress when they are trying to inappropriately move nutrients out of an efficiently working cell. SA would be released when a cell is in no physical danger of survival and might increase with better nutrients or any kind of internal and external environmental conditions above a base line. SA might be a “foundation” indicator, indicating that a cell is in good health without regard to how it is supposed to be functioning (i.e. as a root or as a shoot cell). For example, it might be released after a good rain after a drought, when ABA and the drought action of overall metabolism inhibition, stomate closing, and progressive senescence with plant size shrinkage is no longer warranted.
These Shock/Synchronizer Hormones may also act in low quantities on the metabolism as day-to-day status quo regulators. These actions would involve neither growth nor redirection and its cost of plant shrinkage. ABA and SA levels may rise and fall many times a day. When the levels are low, they may be the equivalent of “moods” in animals or humans or a Circadian Rhythm, alternating between levels of depression and inaction and excitement and action, as the conditions and growth opportunities warrant.
ABA and SA often seem to have counteracting effects (here and here).
10. The Growth Hormones are made primarily in meristematic tissue and the Stress Hormones are made in mature tissue. See figure 1. below or in full size.
|
|
|
Figure 1. Click here for the full Size figure |
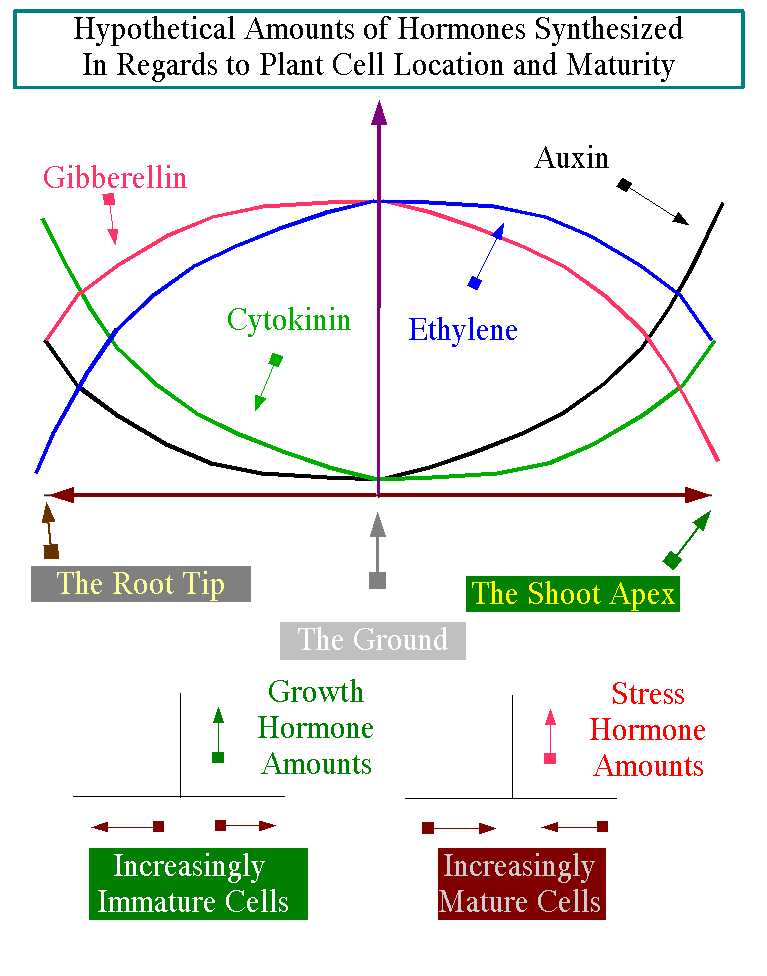
The Shock/Synchronizer Hormones are made in all tissues equally. Although the amount of most hormones made by cells may differ according to their maturity, small amounts of each of these hormone groups are made in all cells under the right conditions. The exception is that perhaps the Shock/Synchronizer Hormones are made in all cells in equal amounts under the right conditions. Alternatively, perhaps under the same conditions, the survival hormone ABA are made in larger amounts in mature cells than in juvenile cells. The suggestion is that juvenile cells can recover from stress more easily than mature cells. Therefore, they have less need of the stress-protecting effects of low levels of ABA and do not need to be sent to the “glue factory” of senescence when they are experiencing high levels of stress (i.e. the plant has “confidence” that they will recover). Perhaps we can even say that plant cells, like all living things, are most susceptible to stress at the beginning and end of their lives. Thus, under the same stress, higher amounts of ABA would be made at the beginning and end of the lives of cells, because the plant would “know” that this was when the cells were most susceptible and least likely to survive stress. Conversely, SA might be more easily made in young cells and mature cells. That is, the highest amounts of SA would be made in cells after they have moved out of the fragile juvenile stage and before they move into senility or close to it.
Getting back to the Growth and Stress Hormones, let’s take Auxin as an example. It is a Growth hormone and primarily a shoot hormone. The largest amounts are made in shoot meristematic cells. Smaller amounts are made in root meristematic cells and also in mature shoot meristematic cells, but there is still a small or very small amount in mature root cells under the right conditions (i.e. under conditions that would induce a shoot meristematic cell to produce it, involving good external conditions and a good level of sugar and oxygen in the older root cells). Looking at ET, it is a Stress hormone and primarily a “shoot hormone” too. It is made when mature shoot cells are experiencing deficiencies in water and minerals, but would also be made when mature root cells are not taking in appropriate amounts of water and minerals. It would also be made in the shoot meristems when they were experiencing deficiencies of minerals and water. Perhaps the ET levels would only rise to levels that would cause hibernation of these meristematic regions, like in the secondary buds. Finally a small or very small amount would be made in meristematic tissue of roots experiencing water and mineral shortage.
 11. Growth Hormones and SA gather all nutrients and perhaps also attract proximate supplies of Growth Hormones and SA while repelling Stress Hormones and ABA. This makes meristematic tissue get involved in positive feedback loops that are responsible for apical dominance, where the better the conditions for dividing cells, the more Growth Hormones and SA are made at the site. This happens in an exponential way. Eventually one meristematic tissue wins out over all the rest, and this becomes the apically dominant meristem. The others go into hibernation. Stress Hormones and ABA repel all nutrients from the site. They may also repel Growth Hormones and SA and attract Stress and ABA.
11. Growth Hormones and SA gather all nutrients and perhaps also attract proximate supplies of Growth Hormones and SA while repelling Stress Hormones and ABA. This makes meristematic tissue get involved in positive feedback loops that are responsible for apical dominance, where the better the conditions for dividing cells, the more Growth Hormones and SA are made at the site. This happens in an exponential way. Eventually one meristematic tissue wins out over all the rest, and this becomes the apically dominant meristem. The others go into hibernation. Stress Hormones and ABA repel all nutrients from the site. They may also repel Growth Hormones and SA and attract Stress and ABA.
12. The Growth Hormones and SA may both need to be at high levels for cell division to take place. The Stress Hormones and ABA may all also need to be “in attendance” before cell death is “signed off on.” If this is true of cell division, the addition of SA may greatly increase the ease of raising calluses from single cells in tissue culture. Where success has been had in the past, the cell lines may have natively synthesized unusually high levels of SA.
13. The ratio of endogenously synthesized Growth Hormones to exogenously available ones will be an important determining factor in morphology. For instance, in a shoot meristem, a cell will store up enough sugars, gases (oxygen and carbon dioxide), minerals (solute concentrations) and water (water pressure) until it “knows” it can reach a mature size. It will then poll its exterior environment and measure the levels of Auxin and CK. If there are high enough levels of these outside the cell, then the cell “knows” that it is a good bet that the supply of sugar, gases, minerals and water will increase. Instead of using the nutrients stored and the resulting hormones synthesized within to make one mature cell, it can risk dividing into two, and trying to create two mature cells.
 14. Increasing levels of Growth Hormones directly inhibit the levels and/or transport of Stress Hormones until a threshold is reached when they directly induce it. This threshold is not tied directly to the Growth Hormone level but is a moving target based on the ratio of the level of the Growth Hormone to the level of the nutrient it is intending to increase. One of the most important reasons for Auxin’s existence and its movement down to the roots is to increase the supply of water and minerals. One of the most important reasons for CK's existence and movement up to the shoots is to increase the supply of sugar and oxygen to the successful new cells in the roots. For example, Auxin will kick off ET production not after Auxin reaches a certain threshold amount, but after it reaches a certain ratio in comparison with the amount of water and or minerals that exist where the Auxin is, in the root or shoot. The point of Auxin is generally to increase the amount of water and minerals with new growth in the roots. If the level of Auxin gets too high in relation to the amount of water and minerals (and the gap is increasing), the plant “knows” that the growth angle is not presently working, and so by promoting the synthesis of Auxin, it tries to “kick start” the mineral and water growth by temporarily inhibiting the synthesis and transport or activating the degradation of itself. ET also has the effect of causing senescence in less efficient mature leaves, thus diminishing the need for water and minerals. The resulting sugar and oxygen are funneled downward to induce a temporary bloom in root growth. The extra minerals and water from this leaf senescence may be sent up to where water is limited, mainly in the shoot meristematic regions that are producing the high levels of Auxin to begin with. This again is a gamble because with the stressing of any nutrients within the plant there are opportunity energy costs.
14. Increasing levels of Growth Hormones directly inhibit the levels and/or transport of Stress Hormones until a threshold is reached when they directly induce it. This threshold is not tied directly to the Growth Hormone level but is a moving target based on the ratio of the level of the Growth Hormone to the level of the nutrient it is intending to increase. One of the most important reasons for Auxin’s existence and its movement down to the roots is to increase the supply of water and minerals. One of the most important reasons for CK's existence and movement up to the shoots is to increase the supply of sugar and oxygen to the successful new cells in the roots. For example, Auxin will kick off ET production not after Auxin reaches a certain threshold amount, but after it reaches a certain ratio in comparison with the amount of water and or minerals that exist where the Auxin is, in the root or shoot. The point of Auxin is generally to increase the amount of water and minerals with new growth in the roots. If the level of Auxin gets too high in relation to the amount of water and minerals (and the gap is increasing), the plant “knows” that the growth angle is not presently working, and so by promoting the synthesis of Auxin, it tries to “kick start” the mineral and water growth by temporarily inhibiting the synthesis and transport or activating the degradation of itself. ET also has the effect of causing senescence in less efficient mature leaves, thus diminishing the need for water and minerals. The resulting sugar and oxygen are funneled downward to induce a temporary bloom in root growth. The extra minerals and water from this leaf senescence may be sent up to where water is limited, mainly in the shoot meristematic regions that are producing the high levels of Auxin to begin with. This again is a gamble because with the stressing of any nutrients within the plant there are opportunity energy costs.
There are four behaviors that Auxin can induce in the root to increase water and mineral supplies. These are:
a. If CK, minerals, and water are high, it can induce cell division in the root meristems and thus increase the supply through new growth without any changes.
b. If CK is low, but minerals and water are high, it can induce new root growth to replace the ineffective root apical meristems and restart mineral and water supply growth.
c. If CK is high but minerals and water are low than this would indicate there is a problem with the functioning of the mature roots. This could be due to inefficient roots but as we will see below it could also be due to healthy roots malfunctioning because of a lack of sugar and oxygen. Either way, the root would want to release at least some ET in order to lower the root nutrient requirements of the shoot and free some sugar and water from cannibalized, less efficient mature leaves. In the case of inefficient roots, the sugar and water would be used to support root growth through cell division. For that, it would also need Auxin. So under this condition, Auxin levels would not be totally suppressed by the new synthesis of ET. On the other hand if the low minerals and water conditions were due to sugar and oxygen starvation of perfectly good roots, then the root would ?? ??
d. If both CK and minerals and water supplies are relatively low in the roots, this means that neither the old roots nor the meristematic roots are working. Auxin will then induce ET. This is done, as mentioned, to lower the mineral and water load, to free both sugar and oxygen for new root growth and to free minerals and water for the shoot. This is “emergency jump start” growth.
In actuality, the first two conditions can also lead to ET emanation if sugar and oxygen levels are low in the roots. When this is true, even if levels of CK and/or minerals and water are high, the roots will not risk cell division or new root initiation because the lack of extra sugar and oxygen is limiting their growth. Auxin’s inducing of ET in the root is tied to its ratio to minerals and water and sugar and oxygen.
To elaborate further, the threshold over which Auxin will induce ET is also tied to the level of GA/BR. Clearly, high levels of GA/BR are an indicator that mature root cells are starving for sugar and oxygen. It is hard to imagine that sugar and oxygen starved roots would be good harvesters of water and minerals from the surrounding soil. The root could monitor just GA/BR to get an idea of the sugar and oxygen needs of the root. Perhaps the root monitors both sugar and oxygen levels and GA/BR levels, with one being the confirmation of the other. The bottom line, however, is the sugar and oxygen level and the root may be able to safely ignore GA/BR levels. Knowing biological systems, however, and their complexity of control, I would not be surprised if Auxin threshold level that would induce ET synthesis is tied to though they would confirm In the end, low levels of sugar and oxygen may lower the threshold for Auxin’s induction of ET. ET inhibits the root senescence promoted by GA/BR. It’s obvious that if this is true it is because it wants to preserve its existing supply of water and minerals
Conversely, increasing levels of Stress Hormones directly inhibit the synthesis and/or transport of the Growth Hormones until again a threshold is reached. Then they encourage the synthesis and/or transport of the Growth Hormones. Again, this threshold is not absolute but is dependent on the ratio of the Stress to the nutrient the Stress are trying to increase. For example, if ET levels have been increasing for a while but minerals and water levels have also been increasing for a while and the ratio between ET and these nutrients has been closing, the plant “knows” the attempt at “jump starting” water and mineral supply has succeeded and ET is no longer needed.
 16. The hormones can be seen as complementary pairs. Auxin’s complement is ET, CK's complement is GA/BR, and SA’s complement is ABA. This is important because Auxin transportation to the root can be seen in part as an attempt to increase water and minerals (even if it is natively synthesized in the root, it leads to cell division or new root growth). If the levels of Auxin rise too high, the plant abandons the attempt temporarily and switches to ET, trying to jump start production. Since Auxin causes cell lengthening and ET causes cell broadening, we can surmise that this kind of thing happens often and provides for balanced growth of the root. ET is perhaps a radical at least temporary change in the root’s strategy for increasing mineral and water supply. If you look back at the chart for ET, there is an entry for a well-known finding that ET induces root hair growth. Root hairs greatly increase the surface area of the root, aiding mineral and water absorption. However, this may make the plant more vulnerable to loss of water during drought. It may also make the plant more susceptible to root predation and disease, because, I hypothesize, the root hair cell is more vulnerable to all these things than the normal wall of the root. So ET represents a major change in strategy for the root. Whereas Auxin causes it to grow down and to make no special arrangements for absorption, ET may cause it to grow out laterally and to make a special of arrangement of growing root hairs. This is all to increase water and mineral levels.
16. The hormones can be seen as complementary pairs. Auxin’s complement is ET, CK's complement is GA/BR, and SA’s complement is ABA. This is important because Auxin transportation to the root can be seen in part as an attempt to increase water and minerals (even if it is natively synthesized in the root, it leads to cell division or new root growth). If the levels of Auxin rise too high, the plant abandons the attempt temporarily and switches to ET, trying to jump start production. Since Auxin causes cell lengthening and ET causes cell broadening, we can surmise that this kind of thing happens often and provides for balanced growth of the root. ET is perhaps a radical at least temporary change in the root’s strategy for increasing mineral and water supply. If you look back at the chart for ET, there is an entry for a well-known finding that ET induces root hair growth. Root hairs greatly increase the surface area of the root, aiding mineral and water absorption. However, this may make the plant more vulnerable to loss of water during drought. It may also make the plant more susceptible to root predation and disease, because, I hypothesize, the root hair cell is more vulnerable to all these things than the normal wall of the root. So ET represents a major change in strategy for the root. Whereas Auxin causes it to grow down and to make no special arrangements for absorption, ET may cause it to grow out laterally and to make a special of arrangement of growing root hairs. This is all to increase water and mineral levels.
Root hair growth may be a normal part of the life of a plant, or it may be a growth gamble not always taken. At any rate, I hypothesize that normally ET, GA/BR, and ABA “rule” the night, in that they are normally released during the night when the plant cannot synthesize sugar or take in as much water and minerals. It is the normal course of events. The lowest levels of Auxin, CK and SA would occur at night and the highest during the day. GA/BR and ET do cause both growth and senescence. That is, at night, when ET is high in the roots, it will be causing lateral growth of the roots, while GA/BR will be causing some senescence of older, less efficient roots. The growth in the roots at night will be balanced, because GA/BR will be lengthening the roots that they are not killing off. In the shoot, ET is doing the converse, pruning older, less efficient leaves while GA is lengthening the good young ones. The good young ones are also broadened by ET at night. I suggest that the plant does the bulk of its self-pruning at night. Also at night, the plant lives off the nutrients it has necessarily stored from the day. In fact, pruning may not be necessary if all parts remain efficient and enough nutrients have been stored from the day to allow for sustenance and even growth.
CK and GA/BR have the same relationship as Auxin and ET. An important reason for transporting CK to the shoot is to increase its sugar and oxygen supply, either by cell division in the meristem in concert with Auxin or by the outgrowth of the secondary buds out of concert with it. If this attempt is unsuccessful CK will induce GA/BR which is a completely different strategy for the shoot. With GA/BR the stem lengthens, in an attempt to move the leaves out of a possible shade and more into the sunlight where more sugar can be synthesized. One would also guess that GA/BR would induce some kind of increase in the efficiency of the leaves, just like root hairs, but again like root hairs, the changed strategy would be more risky. Of course GA/BR does not cause induction of Chlorophyll in seedlings grown in the dark, but in perhaps in low light they might.
 17. When the plant needs to trim or prune parts of itself for reasons other than nutrient deficiencies, such as disease or pestilence, it can and does use the appropriate Stress Hormone as well as ABA. JAs are volatile and may induce a spreading area of senescence as a defense mechanism. For instance, if a leaf gets infected with a disease, the plant will want to limit the spread of the disease, so it will sacrifice cells surrounding the place of infection in order to quarantine the spread. This is done with both ET and ABA. Indeed, this is described in postulates 11 and 12. Even GA/BR will eventually be made as ET and ABA push out nutrients, including sugar and oxygen, from the cells that are being sacrificed. This run-away effect feeds on itself until the cell dies. The spread of self-catalysis after injury or infection is halted perhaps directly by SA and then perhaps indirectly by new Auxin and CK synthesized in response to the influx of nutrients from cannibalized cells from the quarantine area. Indeed wounding, infection, or parasitism may only initially activate ABA directly, and the ET and GA/BR synthesis may be in response to repelling of nutrients that ABA activate. ABA may directly induce SA so that the cannibalization does not go too far. There have been references in the literature to the induction of ABA and SA after wounding or disease. I hypothesize that wounding or infection may only directly activate ABA but this leads to the synthesis of SA. ET and BR/GA would be synthesized indirectly in response to the nutrient repelling action of ABA and Auxin and CK would be indirectly synthesized in response to nutrients being both attracted by SA and pushed out of the cannibalized cells by ABA, ET and GA/BR. Like most biological systems, I hypothesize, there are multiple controls and the notion here of indirect synthesis is incorrect and the plant has more control over the process. Thus, wounding or disease may start just with the release of ABA, but this induces ET and GA/BR at the site and SA, Auxin and CK at some distance from the problem area.
17. When the plant needs to trim or prune parts of itself for reasons other than nutrient deficiencies, such as disease or pestilence, it can and does use the appropriate Stress Hormone as well as ABA. JAs are volatile and may induce a spreading area of senescence as a defense mechanism. For instance, if a leaf gets infected with a disease, the plant will want to limit the spread of the disease, so it will sacrifice cells surrounding the place of infection in order to quarantine the spread. This is done with both ET and ABA. Indeed, this is described in postulates 11 and 12. Even GA/BR will eventually be made as ET and ABA push out nutrients, including sugar and oxygen, from the cells that are being sacrificed. This run-away effect feeds on itself until the cell dies. The spread of self-catalysis after injury or infection is halted perhaps directly by SA and then perhaps indirectly by new Auxin and CK synthesized in response to the influx of nutrients from cannibalized cells from the quarantine area. Indeed wounding, infection, or parasitism may only initially activate ABA directly, and the ET and GA/BR synthesis may be in response to repelling of nutrients that ABA activate. ABA may directly induce SA so that the cannibalization does not go too far. There have been references in the literature to the induction of ABA and SA after wounding or disease. I hypothesize that wounding or infection may only directly activate ABA but this leads to the synthesis of SA. ET and BR/GA would be synthesized indirectly in response to the nutrient repelling action of ABA and Auxin and CK would be indirectly synthesized in response to nutrients being both attracted by SA and pushed out of the cannibalized cells by ABA, ET and GA/BR. Like most biological systems, I hypothesize, there are multiple controls and the notion here of indirect synthesis is incorrect and the plant has more control over the process. Thus, wounding or disease may start just with the release of ABA, but this induces ET and GA/BR at the site and SA, Auxin and CK at some distance from the problem area.
 18. Another “raison d’être” for the growth, and possibly the Stress Hormones too, is to facilitate nutrient transport. Auxin is transported downward, and certainly attracts sugar and oxygen to itself as it travels the phloem subway down to the roots. Incidentally Auxin also attracts minerals and water, so there may be a circulation system in the plant of minerals and water. That goes up the Xylem, but some comes back down in the phloem with Auxin. Similarly, CK is transported up the Xylem, and may take water and minerals coming up with it in the root (although the Xylem is dead wood, so I am not sure about this). Again, similarly to the above, CK might attract sugar and oxygen, as it goes up the hollow tube to the stomata.
18. Another “raison d’être” for the growth, and possibly the Stress Hormones too, is to facilitate nutrient transport. Auxin is transported downward, and certainly attracts sugar and oxygen to itself as it travels the phloem subway down to the roots. Incidentally Auxin also attracts minerals and water, so there may be a circulation system in the plant of minerals and water. That goes up the Xylem, but some comes back down in the phloem with Auxin. Similarly, CK is transported up the Xylem, and may take water and minerals coming up with it in the root (although the Xylem is dead wood, so I am not sure about this). Again, similarly to the above, CK might attract sugar and oxygen, as it goes up the hollow tube to the stomata.